
Lipid: Nhóm Phân Tử Đa Năng Trong Sinh Học và Công Nghệ
11/03/2025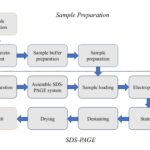
TỔNG QUAN VỀ SDS-PAGE
14/03/2025LIPID HÓA PROTEIN: CƠ CHẾ, PHÁT HIỆN VÀ CÁC BỆNH LÝ LIÊN QUAN
Protein lipid hóa là gì?
Biến đổi sau dịch mã (PTMs) là những thay đổi hóa học xảy ra sau quá trình tổng hợp protein, liên quan đến việc hình thành hoặc phá vỡ các liên kết cộng hóa trị trên khung hoặc chuỗi bên của axit amin. Các PTMs này làm đa dạng hóa cấu trúc và chức năng của protein, góp phần tạo nên sự phong phú của sinh vật.
Lipid hóa là một loại PTM phổ biến ở tế bào nhân thực, trong đó các nhóm lipid hoặc các nhóm giống lipid được gắn vào protein. Quá trình này làm tăng tính kỵ nước của protein, cho phép chúng gắn vào màng tế bào hoặc tham gia vào các quá trình sinh học quan trọng khác.
Lipid hóa giúp protein tương tác với các thành phần khác trong tế bào, tham gia vào các con đường truyền tín hiệu và định vị protein đến các vị trí cụ thể, đặc biệt là màng tế bào. Ví dụ, protein G, đóng vai trò trung tâm trong nhiều con đường truyền tín hiệu, được lipid hóa để gắn vào mặt trong của màng sinh chất. Điều này cho phép chúng tương tác với các thụ thể trên bề mặt tế bào và kích hoạt các con đường truyền tín hiệu. Các tín hiệu này điều chỉnh nhiều quá trình quan trọng như dẫn truyền thần kinh, đáp ứng hormone và phản ứng miễn dịch.
Có bao nhiêu loại biến đổi?
1. S-acyl hóa
S-acyl hóa là một quá trình có ở hầu hết các sinh vật nhân thực (từ nấm men đến con người) và được kiểm soát bởi một nhóm enzyme đặc biệt. Mặc dù vi khuẩn không có S-acyl hóa, nhưng một số virus có thể lợi dụng hệ thống enzyme của tế bào chủ để thực hiện quá trình này. Không giống như các dạng biến đổi lipid khác (thường là vĩnh viễn), S-acyl hóa có thể bị loại bỏ hoặc gắn lại nhiều lần nhờ liên kết thioester dễ bị phá vỡ trong môi trường nội bào. Điều này cho phép protein liên tục điều chỉnh vị trí và chức năng của chúng trong tế bào.
Khi nói đến S-acyl hóa, người ta thường nhắc đến palmitoyl hóa – tức là việc gắn palmitate (C16:0) vào protein. Tuy nhiên, các axit béo khác như stearate (C18:0) và oleate (C18:1) cũng có thể được gắn vào protein theo cách này. Các protein bị S-acyl hóa rất đa dạng, từ protein xuyên màng đến protein bám màng.
- Nhóm protein bám màng (Peripheral membrane proteins): Chúng thường cần có hai tín hiệu lipid để gắn chặt vào màng, chẳng hạn như:
- N-myristoyl hóa + S-acyl hóa (ví dụ: Src kinases)
- Prenyl hóa + S-acyl hóa (ví dụ: protein Ras)
- Chỉ có S-acyl hóa (ví dụ: SNAP25, một protein quan trọng trong việc xuất bào).
- Nhóm protein xuyên màng (Transmembrane proteins): Gồm nhiều loại khác nhau như kênh ion, thụ thể và protein vận chuyển. S-acyl hóa giúp điều chỉnh vị trí và chức năng của chúng trên màng tế bào.
Các enzyme xúc tác
Các nghiên cứu độc lập trên nấm men Saccharomyces cerevisiae đã xác định những enzyme kiểm soát quá trình s-acyl hóa được đặt tên là Erf2p và Akr1p. Khi phân tích trình tự của chúng, các nhà khoa học nhận thấy rằng cả hai đều có một vùng kẽm ngón tay (zinc-finger domain) dài 51 axit amin với một motif “DHHC” đặc trưng. DHHC là viết tắt của bốn axit amin đặc trưng trong vùng xúc tác của enzyme: Aspartate (D), Histidine (H), Histidine (H), Cysteine (C).
Vùng DHHC nằm trong miền zinc-finger của enzyme và đóng vai trò quan trọng trong quá trình gắn nhóm acyl (thường là palmitate) vào protein đích.
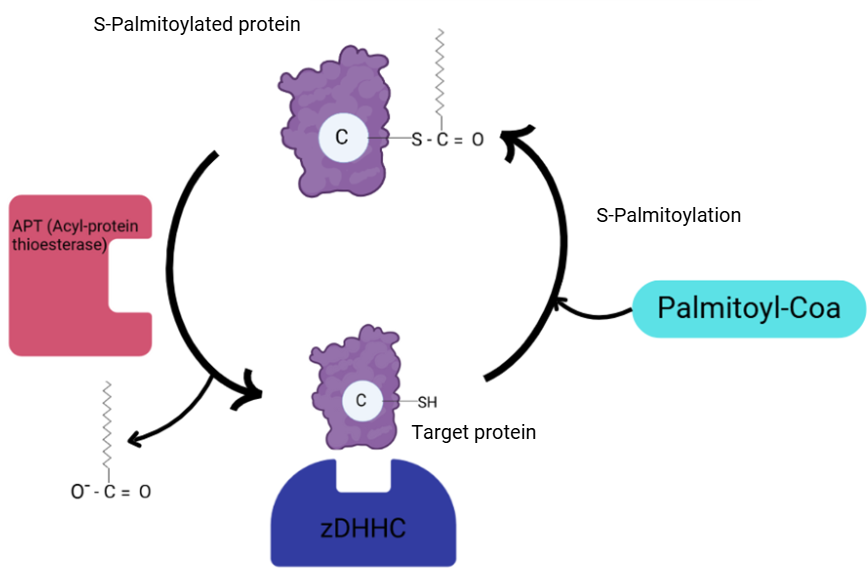
Hình 1. Quá trình palmitoyl hóa protein và loại bỏ nhóm palmitoyl khỏi protein.
(Lộc, Research Intern, Hoan Vu Biomolecules., JSC)
Tác động điều hòa của S-acyl hóa
Liên kết màng của protein ngoại vi:
- S-acyl hóa giúp protein ngoại vi bám chặt vào màng tế bào, ngay cả khi protein đã có sẵn nhóm lipid khác như myristoyl hoặc prenyl.
- Nếu protein chỉ có một nhóm lipid, liên kết với màng sẽ yếu và tạm thời. Khi được S-acyl hóa, protein sẽ gắn chặt hơn vào màng, một quá trình gọi là “bẫy động học” (kinetic trapping).
- Ví dụ: Nếu Ras protein mất vị trí S-acyl hóa, nó sẽ liên kết yếu với màng. Ngược lại, nếu mất farnesylation, Ras không thể bị S-acyl hóa và không bám vào màng được.
Định vị protein vào các vi miền màng:
- Màng tế bào chứa nhiều loại lipid khác nhau, có thể tạo thành các vi miền đặc biệt (lipid rafts) giàu cholesterol và phospholipid bão hòa.
- S-acyl hóa giúp protein tập trung vào các vi miền này, ảnh hưởng đến vị trí và chức năng của chúng trong màng tế bào.
- Ví dụ:
- SNAP25: Một protein quan trọng trong xuất bào, cần S-acyl hóa để hoạt động hiệu quả.
- Rac1: Liên quan đến tổ chức khung xương tế bào và cấu trúc màng, cần S-acyl hóa để điều hòa chức năng.
Định hướng protein đến các bào quan nội bào:
- Không chỉ màng tế bào, các bào quan như lưới nội chất (ER) cũng có vi miền riêng biệt.
- S-acyl hóa giúp protein được định vị chính xác trong các vùng này. Ví dụ:
- Calnexin: Một protein hỗ trợ gấp nếp glycoprotein trong ER, cần S-acyl hóa để tập trung vào vùng ER thô gần nhân.
- TMX và heme oxygenase-1: Cần S-acyl hóa để đến màng liên kết với ty thể (MAM – mitochondrial-associated membrane).
2. N-myristoyl hóa
Gắn nhóm myristoyl (C14:0, một acid béo 14 carbon) vào glycine ở đầu N của protein thông qua liên kết amide. Quá trình này có thể diễn ra ngay trong lúc dịch mã (co-translational) hoặc sau khi protein được tổng hợp (post-translational).
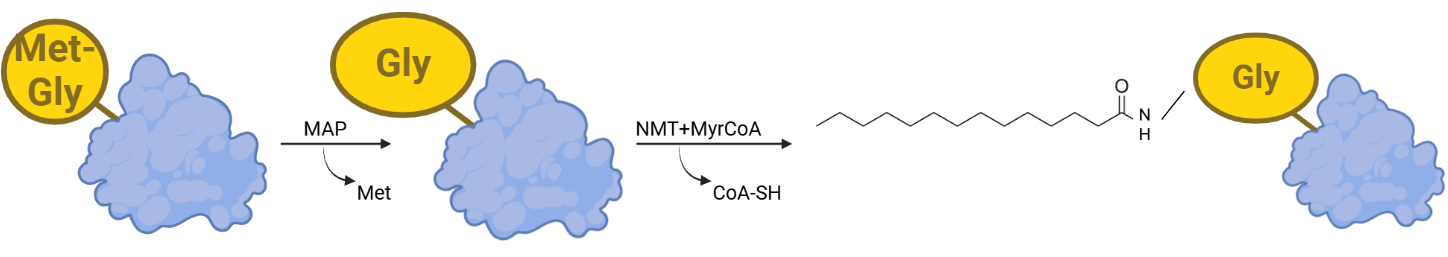
Hình 2. Quá trình myristoyl hóa protein.
(Lộc, Research Intern, Hoan Vu Biomolecules., JSC)
Các enzyme xúc tác
Quá trình được xúc tác bởi enzyme N-myristoyltransferase (NMT). Điều này giúp protein gắn vào màng tế bào và thực hiện nhiều chức năng quan trọng trong cơ thể. NMT có 2 dạng: NMT1 và NMT2.
- NMT1 quan trọng trong sự phát triển phôi và tế bào ung thư.
- NMT2 có chức năng điều hòa bổ trợ.
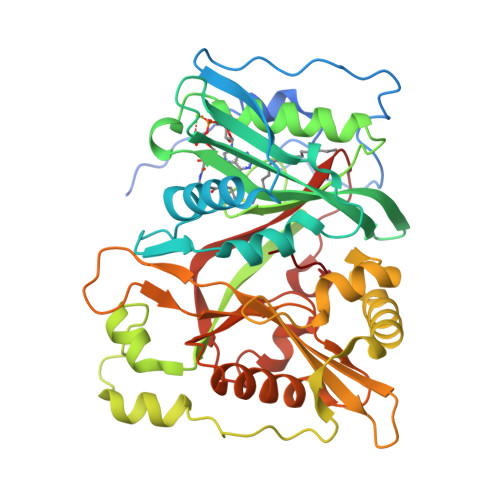
Hình 3. NMT 2
(PDB ID: 4C2X)
Tác động điều hòa của myristoyl hóa
Hướng protein đến màng tế bào. Tuy nhiên, chỉ có myristoyl hóa thôi thì chưa đủ để protein bám vào màng – nó cần thêm một tín hiệu khác, như:
- Các biến đổi lipid gần đó (ví dụ: palmitoyl hóa trên cysteine hoặc prenyl hóa trên cysteine).
- Sự có mặt của các nhóm amino acid mang điện tích dương.
Sự kết hợp này tạo ra một cơ chế gọi là “công tắc myristoyl”, trong đó việc protein bám vào hay tách ra khỏi màng có thể được kiểm soát bằng quá trình phosphoryl hóa hoặc sự liên kết với các phân tử như GTP và Ca²⁺.
Các dạng lipid hóa như myristoyl hóa và palmitoyl hóa giúp protein bám vào màng tế bào, từ đó tạo nền tảng cho các tín hiệu nội bào. Các protein được cải biến gắn thêm chuỗi myristol thường gặp là các tiểu đơn vị α của protein G thể dị tam (heterotrimeric G proteins), các protein G thể đơn (monomeric G protein) và tyrosine kinase có mặt ở tế bào chất.
Ổn định protein và giúp chúng tương tác với các phân tử khác: Myristoyl hóa không chỉ giúp protein gắn màng mà còn ảnh hưởng đến cách chúng tương tác với các protein khác. Ví dụ: CAP-23/NAP-22, một protein trong não, cần myristoyl hóa để tương tác hiệu quả với calmodulin – một protein điều hòa tín hiệu canxi.
Kiểm soát hoạt động của enzyme: Một ví dụ điển hình là cơ chế “công tắc myristoyl” giúp kiểm soát hoạt động của c-Abl tyrosine kinase.
- Bình thường, c-Abl ở trạng thái không hoạt động.
- Khi có myristoyl hóa, cấu trúc protein thay đổi, giữ nó trong trạng thái “tắt”.
- Nếu mất myristoyl hóa, c-Abl sẽ hoạt động mạnh hơn, có thể gây ra sự phát triển không kiểm soát của tế bào (liên quan đến ung thư).
Điều chỉnh vị trí và chức năng của protein trong apoptosis: Diễn ra khi enzyme caspase cắt protein trong quá trình apoptosis (chết tế bào theo chương trình). Việc thêm nhóm myristoyl giúp protein có chức năng đặc biệt, như điều chỉnh quá trình chết tế bào. Ví dụ: BID, một protein thúc đẩy apoptosis, bị caspase-8 cắt thành hai mảnh. Mảnh C-terminal của BID sau đó được myristoyl hóa, giúp nó gắn vào màng ty thể, kích hoạt quá trình chết tế bào.
3. S-prenyl hóa
Prenyl hóa (prenyl hóa) chỉ xuất hiện trong tế bào nhân thực (eukaryote), và hầu hết các protein đã biết có prenyl hóa đều thuộc về tế bào nhân thực. Tuy nhiên, một số vi khuẩn và virus có thể lợi dụng hệ thống prenyl hóa của vật chủ để giúp chúng xâm nhập và tồn tại bên trong tế bào vật chủ. Ví dụ như protein Salmonella-induced filament A (từ vi khuẩn Salmonella typhimurium) bị geranylgeranyl hóa bởi GGT-1 của vật chủ.
Quá rình gắn nhóm isoprenoid (farnesyl hoặc geranylgeranyl) vào cysteine ở vị trí thứ 4 tính từ đầu C của chuỗi polypeptide thông qua liên kết thioether, giúp tăng độ bền cho protein.
Có hai loại chính của prenyl hóa:
- Farnesyl hóa: gắn nhóm farnesyl (có 3 đơn vị isopren). Ví dụ: Lamin B cũng cần farnesyl hóa để hình thành màng nhân ổn định trong phân bào. Ở chuột, thiếu farnesylation ở lamin B1 gây tử vong ngay sau sinh.
- Geranylgeranyl hóa: gắn nhóm geranylgeranyl (có 4 đơn vị isopren). Ví dụ: Rab proteins, được geranylgeranyl hóa kép bởi GGTase-II, giúp vận chuyển các thành phần trong con đường Notch. Nếu Rab bị định vị sai, tín hiệu Notch sẽ bị gián đoạn, gây rối loạn phát triển.
Đa số protein bị prenyl hóa theo kiểu geranylgeranyl hóa. Một vài protein được prenyl hóa. Một số protein có thể bị cả hai loại prenyl hóa: Ví dụ như K-Ras, N-Ras và RhoB có thể vừa bị farnesyl hóa (FT) vừa bị geranylgeranyl hóa (GGT-1).
Protein Ras trải qua quá trình farnesyl hóa hoặc geranylgeranyl hóa để gắn vào màng tế bào.
Các enzyme xúc tác
- FTase (Farnesyltransferase): Gắn nhóm farnesyl (15C) từ phân tử farnesyl diphosphate (FPP) vào protein có motif CAAX ở đầu C.
- GGTase-I (Geranylgeranyltransferase-I): Gắn nhóm geranylgeranyl (20C) vào protein có motif CAAX.
- GGTase-II (Rab Geranylgeranyltransferase): Có thể gắn một hoặc hai nhóm geranylgeranyl vào protein chứa motif CC, CXC, CCX, CCXX hoặc CCXXX.
- GGTase-III: Chịu trách nhiệm gắn hai nhóm geranylgeranyl vào một số protein đặc biệt như FBXL2 và Ykt6.
Lưu ý:
Một chuỗi amino acid đặc trưng ở đầu C tận (C-terminal), gọi là motif CaaX.
C = Cysteine
A = Aliphatic amino acid (Leu, Ile, Val, Ala…)
A = Aliphatic amino acid
X = Bất kỳ amino acid nào
CC, CXC, CCX, CCXX, CCXXX Motif (đặc trưng của protein Rab)
Motif CAAX đóng vai trò quan trọng trong việc nhận diện enzyme:
Nếu X = Ala, Met, Ser, Glu, protein sẽ được farnesyl hóa.
Nếu X = Leu, Met, Phe, Ile, Val, protein sẽ được geranylgeranyl hóa.
Protein chỉ trải qua geranylgeranyl hóa (GG hóa) bởi GGTase-II (RabGGTase)
FTase và GGTase-I có cùng tiểu đơn vị α (FNTA hoặc PTAR2) nhưng khác nhau ở tiểu đơn vị β, giúp quyết định tính đặc hiệu của enzyme.
Tác động điều hòa của prenyl hóa
- Giúp protein bám lên màng tế bào thông qua farnesyl hóa, geranylgeranyl hóa và các bước hậu xử lý như palmitoyl hóa, thủy phân, methyl hóa.
- Ảnh hưởng đến tương tác protein.
- Giúp ổn định protein, nhưng cũng có thể thúc đẩy sự phân hủy protein tùy theo bối cảnh.
- Có vai trò quan trọng trong truyền tín hiệu, đặc biệt là trong điều hòa hệ miễn dịch và kiểm soát sự phát triển tế bào.
4. Neo GPI
Gắn “neo” glycosylphosphatidylinositol (GPI): GPI là một loại mỏ neo lipid giúp gắn nhiều loại protein lên bề mặt tế bào. Kiểu biến đổi protein bằng cách gắn thêm “neo” GPI có tính thuận nghịch, vì protein được neo có thể tách ra khỏi màng tế bào nhờ xúc tác của phospholipase. Trong một số trường hợp, quá trình giải phóng protein (enzym) ra khỏi màng có thể dẫn đến sự hoạt hóa enzym.
GPI phổ biến ở sinh vật nhân thực và một số vi khuẩn cổ. Ở người, có ít nhất 150 protein GPI tham gia vào nhiều chức năng quan trọng như thụ thể, enzyme, phân tử kết dính và vận chuyển. Các protein được neo bởi GPI thường tham gia vào các quá trình hấp thu dinh dưỡng, kết dính tế bào và truyền tín hiệu qua màng tế bào.

Hình 4. Vị trí neo GPI
(Lộc, Research Intern, Hoan Vu Biomolecules., JSC)
Tổng quan về quá trình sinh tổng hợp và vận chuyển GPI
GPI (Glycosylphosphatidylinositol) là một loại neo lipid giúp gắn các protein lên màng tế bào. Quá trình tổng hợp GPI diễn ra trong ba giai đoạn chính:
- Tổng hợp tiền chất GPI trong màng lưới nội chất (ER).
- Gắn GPI vào protein mới tổng hợp trong lòng ER, đồng thời cắt bỏ tín hiệu peptid C tận.
- Chỉnh sửa lipid và carbohydrate trong ER và sau khi vận chuyển đến bộ máy Golgi.
Giai đoạn 1: Tổng hợp tiền chất GPI
- GPI được tổng hợp từ UDP-GlcNAc (uridine diphosphate N-acetylglucosamine) và PI (phosphatidylinositol), tạo thành GlcNAc-PI.
- GlcNAc-PI sau đó bị khử acetyl tạo thành GlcN-PI ở phía bào tương của ER.
- Từ đây, quá trình sinh tổng hợp GPI có sự khác biệt giữa các loài:
- Ở Trypanosoma brucei, GlcN-PI được gắn thêm nhóm inositol acyl sau khi thêm mannose đầu tiên.
- Ở nấm men (Saccharomyces cerevisiae) và tế bào động vật có vú, quá trình này diễn ra theo thứ tự ngược lại: trước tiên là inositol acyl hóa, sau đó mới thêm mannose.
Giai đoạn 2: Gắn GPI vào protein
- GPI được gắn vào protein thông qua enzyme GPI transamidase. Enzyme này nhận diện trình tự tín hiệu GPI (GPI-addition signal peptide, viết tắt là GPIsp) ở đầu C tận của protein.
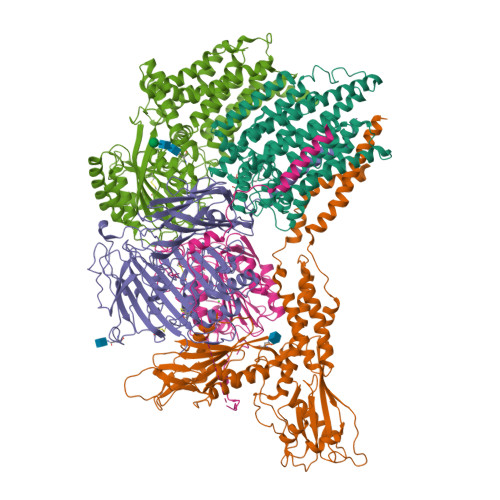
Hình 5. Cấu trúc của enzyme GPI transamidase ở người
(PDB ID: 7W72)
- Trình tự GPIsp có ba đặc điểm chính:
- Ba amino acid nhỏ (Ala, Asn, Asp, Cys, Gly hoặc Ser) tại vị trí ω, ω+1 và ω+2.
- Một vùng phân cực gồm 5–10 amino acid.
- Một vùng kỵ nước dài 15–20 amino acid.
- Enzyme GPI transamidase thay thế đoạn tín hiệu này bằng một neo GPI, giúp protein bám vào màng tế bào.
Giai đoạn 3: Chỉnh sửa GPI
Sau khi gắn vào protein, GPI tiếp tục được biến đổi để phù hợp với chức năng của từng loại tế bào và loài sinh vật:
- Ở nấm men, quá trình này xảy ra sau khi gắn vào protein, với hai bước:
- Axit béo không bão hòa (C18:1) ở vị trí sn-2 được thay thế bằng C26:0.
- Diacylglycerol trong GPI được thay thế bằng ceramide.
- Ở tế bào động vật có vú, quá trình này phức tạp hơn, liên quan đến sự thay đổi từ diacyl-PI sang diradyl-PI (hỗn hợp của 1-alkyl-2-acyl-PI và diacyl-PI).
Vận chuyển GPI-APs (GPI-anchored proteins)
- Các protein gắn GPI được vận chuyển từ ER đến Golgi nhờ các túi vận chuyển được phủ protein COPII.
- Quá trình này yêu cầu một thụ thể xuyên màng là p24, giúp liên kết GPI với các thành phần COPII trên màng bào tương.
Phương pháp phát hiện
Trong nghiên cứu sinh hóa và sinh học tế bào, việc xác định sự gắn lipid vào protein đóng vai trò quan trọng trong việc làm sáng tỏ các cơ chế tín hiệu và điều hòa chức năng của protein. Các phương pháp phát hiện sự biến đổi lipid trên protein đã phát triển đáng kể, từ các kỹ thuật cổ điển sử dụng phóng xạ đến những phương pháp hiện đại không sử dụng phóng xạ, giúp nâng cao độ nhạy và an toàn trong nghiên cứu.
1. S-Acyl hóa trên protein
Bảng 1. So sánh 2 phương pháp phát hiện protein bị S-acyl hóa.
(Lộc, Research Intern, Hoan Vu Biomolecules., JSC)
| Phương pháp | Nguyên lý | Ưu điểm | Nhược điểm |
| Đánh dấu lipid bằng acid béo chứa nhóm phản ứng (17-ODYA, Alk-C16) | Sử dụng acid béo chứa nhóm phản ứng, sau đó gắn chất huỳnh quang hoặc biotin bằng click chemistry. | Chính xác, theo dõi động học S-Acyl hóa. | Chỉ áp dụng trên tế bào sống, không phân biệt được loại lipid gắn vào protein. |
| Acyl-Biotin Exchange (ABE) và Acyl-RAC | Cắt liên kết thioester bằng hydroxylamine, gắn biotin hoặc thu giữ bằng resin, phát hiện bằng Western blot hoặc khối phổ. | Không cần nuôi tế bào với lipid đánh dấu, áp dụng trên mẫu mô. | Không xác định được loại lipid gắn vào cysteine. |
2. N-myristoyl hóa
Có nhiều phương pháp được phát triển để phát hiện N-myristoyl hóa trong tế bào sống (in vivo) và trong ống nghiệm (in vitro).
Bảng 2. So sáng giữa phương pháp cổ điển và phương pháp hiện đại nhằm phát hiện protein bị myristoyl hóa.
(Lộc, Research Intern, Hoan Vu Biomolecules., JSC)
| Phương pháp | Nguyên lý | Ưu điểm | Nhược điểm |
| Cổ điển (Sử dụng phóng xạ) | Sử dụng axit myristic được đánh dấu phóng xạ ([³H]-myristic acid hoặc [¹²⁵I]-myristic acid). Protein mục tiêu được kết tủa bằng kháng thể và phát hiện trên phim phóng xạ. | Xác định chính xác protein đã gắn myristic. | Nhạy kém, tốn thời gian. Phải xử lý chất phóng xạ, nguy hiểm và khó kiểm soát. |
| Hiện đại (Không dùng phóng xạ) | Sử dụng các chất tương tự axit myristic có nhóm ω-azido hoặc ω-alkynyl. Protein bị biến đổi được đánh dấu bằng huỳnh quang hoặc biotin thông qua phản ứng hóa học đặc hiệu (Staudinger ligation hoặc Huisgen cycloaddition). Protein có đánh dấu được phát hiện bằng điện di gel SDS-PAGE hoặc western blot. | An toàn, không phóng xạ. Nhạy hơn, dễ thực hiện trong tế bào sống. Ứng dụng rộng rãi trong nhiều nghiên cứu. | Phụ thuộc vào phản ứng hóa học đặc hiệu. Có thể tạo nhiễu với một số lipid khác. |

Hình 6. Phương pháp hiện đại (không dùng phóng xạ)
(Lộc, Research Intern, Hoan Vu Biomolecules., JSC)
3. Prenyl Hóa
Bảng 3. So sánh 2 phương pháp phát hiện protein bị prenyl hóa.
(Lộc, Research Intern, Hoan Vu Biomolecules., JSC)
| Phương pháp | Nguyên lý | Ưu điểm | Nhược điểm |
| Dò đồng vị phóng xạ | Sử dụng các hợp chất phóng xạ ([³H]-DATFP-FPP, [³²P]-DATFP-GPP) để gắn vào enzyme farnesyltransferase (FT), GGT-1 và RGGT, sau đó phân tích sự gắn kết. | Chính xác trong nghiên cứu hoạt tính enzyme. | Độ nhạy thấp, cần thời gian phát hiện dài (vài ngày). Không có nhãn trực tiếp để xác định protein mục tiêu. Nguy hiểm khi xử lý chất phóng xạ. |
| Dò huỳnh quang | Sử dụng các hợp chất huỳnh quang (NBD-FPP, N-methylanthraniloyl isoprenoid diphosphate) để đánh dấu protein bị prenyl hóa, theo dõi quá trình này trong tế bào sống. | Dễ dàng theo dõi trực tiếp bằng kính hiển vi huỳnh quang. Giúp sàng lọc nhanh các chất ức chế enzyme prenyltransferase. | Một số dẫn xuất huỳnh quang có thể ảnh hưởng đến quá trình sinh học tự nhiên. Không phải tất cả protein đều gắn dò huỳnh quang hiệu quả. |

Hình 7. Dò huỳnh quang để theo dõi prenyl hóa
(Lộc, Research Intern, Hoan Vu Biomolecules., JSC)
4. Neo GPI
Bảng 4. So sánh 2 phương pháp phát hiện protein gắn neo GPI.
(Lộc, Research Intern, Hoan Vu Biomolecules., JSC)
| Phương pháp | Nguyên lý | Ưu điểm | Nhược điểm |
| Phương pháp tách lớp bằng chất tẩy rửa | Sử dụng chất tẩy rửa Triton X-114 để tách lớp protein có GPI khỏi các protein khác. Phát hiện bằng Western blot hoặc kháng thể đặc hiệu. | Dễ thực hiện, không cần enzyme hoặc hóa chất mạnh. Có thể kết hợp với phương pháp đánh dấu phóng xạ để tăng độ chính xác. | Độ chính xác không cao bằng các phương pháp cắt trực tiếp. Không phân biệt được tất cả protein có GPI vì một số protein có thể bị nhiễu. |
| Phương pháp đánh dấu phóng xạ | Dùng các hợp chất phóng xạ (myo-inositol, ethanolamine, glucosamine, mannose, axit béo) để đánh dấu protein có GPI. Phát hiện bằng sắc ký hoặc Western blot. | Rất nhạy, có thể phát hiện protein ở nồng độ thấp. Xác định chính xác thành phần GPI của protein. | Cần thiết bị phóng xạ chuyên dụng, phức tạp khi xử lý. Độc hại, yêu cầu an toàn sinh học cao. |
Bệnh lý
1. S-acyl hóa
Vận chuyển cholesterol qua màng tế bào
Được thực hiện bởi hai protein ABCA1 và ABCG1 thuộc họ ATP-binding cassette (ABC transporters), giúp đưa cholesterol ra khỏi tế bào để tạo lipoprotein mật độ cao (HDL). S-acyl hóa đóng vai trò quan trọng trong quá trình này.
ABCA1 cần S-acyl hóa để di chuyển từ lưới nội chất (ER) ra màng tế bào, nếu không có S-acyl hóa, cholesterol không thể xuất ra khỏi tế bào. S-acyl hóa giúp protein vận chuyển cholesterol hoạt động bình thường, và rối loạn quá trình này có thể liên quan đến các bệnh như xơ vữa động mạch, thoái hóa điểm vàng hoặc ung thư.
Vai trò của S-acyl hóa trong kênh CFTR (có liên quan đến bệnh xơ nang – cystic fibrosis)
Kênh CFTR kiểm soát sự vận chuyển chloride và bicarbonate, rất quan trọng trong việc duy trì chất nhầy ở phổi. Đột biến F508del làm mất một axit amin trong CFTR, khiến protein bị gấp cuộn sai và bị phân hủy trước khi đến được màng tế bào.
Tuy nhiên, S-acyl hóa giúp CFTR, kể cả dạng đột biến F508del, được ổn định và vận chuyển tốt hơn từ lưới nội chất đến Golgi. Điều này mở ra hướng mới trong điều trị xơ nang bằng cách kết hợp S-acyl hóa với các thuốc chỉnh sửa CFTR hiện có.
2. N- myristoyl hóa
NMT là mục tiêu điều trị nấm và ký sinh trùng
Nhiều nghiên cứu cho thấy NMT là mục tiêu tiềm năng để phát triển thuốc chống nấm và chống ký sinh trùng vì:
- Nó cần thiết cho sự phát triển và sống sót của nấm và ký sinh trùng.
- Vị trí liên kết peptide của NMT khác nhau giữa các loài, giúp thiết kế thuốc chọn lọc chỉ nhắm vào vi sinh vật mà không ảnh hưởng đến tế bào người.
Các chất ức chế NMT đã được phát triển để chống lại. Ví dụ: Plasmodium falciparum (ký sinh trùng sốt rét): Khi NMT bị ức chế, tế bào ký sinh trùng không thể duy trì cấu trúc nội bào và sẽ chết.
Virus và vi khuẩn lợi dụng hệ thống N-myristoyl hóa của vật chủ
Một số virus và vi khuẩn sử dụng hệ thống N-myristoyl hóa của tế bào chủ để lây nhiễm. Ví dụ: Virus HIV-1 cần N-myristoyl hóa của protein Gag để hình thành và lắp ráp hạt virus.
Ngoài ra, vi khuẩn Shigella flexneri, tác nhân gây tiêu chảy, có thể gây rối loạn hệ thống N-myristoyl hóa của tế bào chủ thông qua enzyme IpaJ. Enzyme này cắt đứt liên kết N-myristoyl hóa, làm gián đoạn các tín hiệu quan trọng trong tế bào, giúp vi khuẩn xâm nhập và phát triển.
Bên cạnh đó, NMT cũng là mục tiêu cho liệu pháp chống ung thư vì protein bị N-myristoyl hóa tham gia vào con đường tín hiệu và quá trình apoptosis. Khi xảy ra đột biến, chúng có thể kích hoạt sự phát triển không kiểm soát của tế bào ung thư.
3. Prenyl Hóa
Một số vi khuẩn và virus có thể lợi dụng hệ thống prenyl hóa của vật chủ để giúp chúng xâm nhập và tồn tại bên trong tế bào vật chủ:
- Vi khuẩn: Protein Ankyrin B (ANKB) của Legionella pneumophila bị farnesyl hóa bởi FT của vật chủ, giúp nó bám vào màng tế bào chủ và hỗ trợ vi khuẩn nhân lên bên trong.
- Virus: Protein kháng nguyên lớn của virus viêm gan D (HDV) có motif CaaX, giúp nó được prenyl hóa để tạo thành virus hoàn chỉnh. Thuốc ức chế prenyl hóa có thể ngăn chặn sự hình thành virus viêm gan D. Một nghiên cứu lâm sàng giai đoạn 2A đã cho thấy lonafarnib (một thuốc ức chế prenyl hóa) có thể làm giảm lượng virus trong cơ thể người bệnh.
4. GPI-APs (glycosylphosphatidylinositol-anchored proteins)
Một số sinh vật kí sinh ở người có các enzym đính trên bề mặt tế bào thông qua “neo” GPI. Các enzym này thường ở trạng thái bất hoạt nhưng sẽ được kích hoạt bởi phospholipase vật chủ. Như trường hợp ký sinh trùng Trypanosoma brucei (gây bệnh ngủ châu Phi). Trypanosoma brucei có một lớp vỏ bảo vệ gọi là VSG (Variant Surface Glycoproteins), được neo GPI. Khi phospholipase C của vật chủ cắt bỏ neo GPI, VSG bị giải phóng, giúp Trypanosoma thay đổi kháng nguyên để trốn thoát hệ miễn dịch.
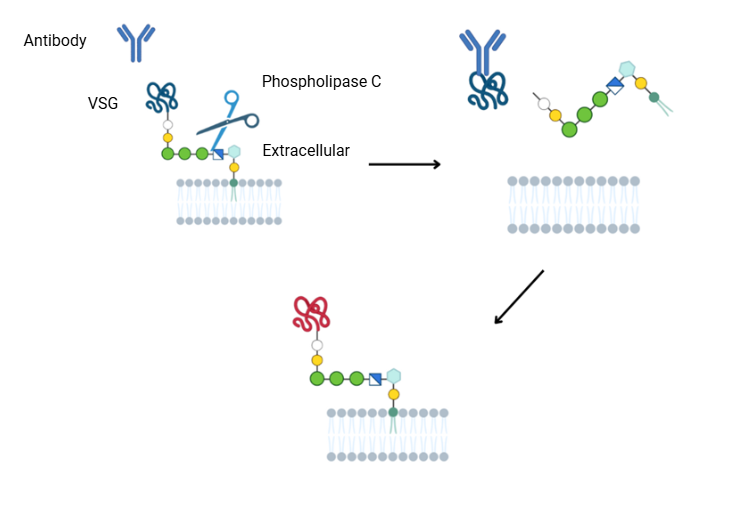
Hình 8. Cách để Trypanosoma brucei (gây bệnh ngủ châu Phi) trốn thoát hệ miễn dịch.
(Lộc, Research Intern, Hoan Vu Biomolecules., JSC)
TLTK
[1] Jiang, H., Zhang, X., Chen, X., Aramsangtienchai, P., Tong, Z., & Lin, H. (2018). Protein Lipidation: Occurrence, Mechanisms, Biological Functions, and Enabling Technologies. Chemical reviews, 118(3), 919–988. https://doi.org/10.1021/acs.chemrev.6b00750
[2] Zhong, Q., Xiao, X., Qiu, Y., Xu, Z., Chen, C., Chong, B., Zhao, X., Hai, S., Li, S., An, Z., & Dai, L. (2023). Protein posttranslational modifications in health and diseases: Functions, regulatory mechanisms, and therapeutic implications. MedComm, 4(3), e261. https://doi.org/10.1002/mco2.261
[3] Pisanti, S., Rimondi, E., Pozza, E., Melloni, E., Zauli, E., Bifulco, M., Martinelli, R., & Marcuzzi, A. (2022). Prenyl hóa Defects and Oxidative Stress Trigger the Main Consequences of Neuroinflammation Linked to Mevalonate Pathway Deregulation. International journal of environmental research and public health, 19(15), 9061. https://doi.org/10.3390/ijerph19159061
[4] Chamberlain, L. H., & Shipston, M. J. (2015). The physiology of protein S-acyl hóa. Physiological reviews, 95(2), 341–376. https://doi.org/10.1152/physrev.00032.2014
[5] Wang, B., Dai, T., Sun, W. et al. Protein N-myristoyl hóa: functions and mechanisms in control of innate immunity. Cell Mol Immunol 18, 878–888 (2021). https://doi.org/10.1038/s41423-021-00663-2
[6] Yuan, Y., Li, P., Li, J. et al. Protein lipidation in health and disease: molecular basis, physiological function and pathological implication. Sig Transduct Target Ther 9, 60 (2024). https://doi.org/10.1038/s41392-024-01759-7
[7] Qi Zhang, Morihisa Fujita, Why nature evolved GPI-anchored proteins: unique structure characteristics enable versatile cell surface functions, Glycobiology, Volume 34, Issue 12, December 2024, cwae089, https://doi.org/10.1093/glycob/cwae089
[8] Komath SS, Fujita M, Hart GW, et al. Glycosylphosphatidylinositol Anchors. In: Varki A, Cummings RD, Esko JD, et al., editors. Essentials of Glycobiology [Internet]. 4th edition. Cold Spring Harbor (NY): Cold Spring Harbor Laboratory Press; 2022. Chapter 12. Available from: https://www.ncbi.nlm.nih.gov/books/NBK579963/ doi: 10.1101/glycobiology.4e.12
[9] Nguyễn Thị Hồng Thương, Nguyễn Tiến Thắng. (2020). Giáo trình hóa học protein. TP. Hồ Chí Minh : Đại học Quốc gia TP. Hồ Chí Minh.
BÀI VIẾT MỚI >>
- Các phương pháp phân tích định lượng, định tính
 Các phương pháp phân tích định lượng, định tính Giá trên đã bao gồm thuế phí.
Các phương pháp phân tích định lượng, định tính Giá trên đã bao gồm thuế phí. - Các phương pháp phân tích Sinh Hóa Lý
- Tổng quan về ELISA
 ELISA (Enzyme-linked immunosorbent assay) là kỹ thuật xét nghiệm miễn dịch sử dụng enzyme để phát hiện và định lượng kháng nguyên hoặc kháng thể. Có […]
ELISA (Enzyme-linked immunosorbent assay) là kỹ thuật xét nghiệm miễn dịch sử dụng enzyme để phát hiện và định lượng kháng nguyên hoặc kháng thể. Có […] - Các phương pháp chiết tách protein trong nghiên cứu proteomics và ứng dụng
 Các tiến bộ trong công nghệ proteomics không thể khắc phục được các vấn đề trong chuẩn bị mẫu. Các bước như đồng nhất hóa mô, […]
Các tiến bộ trong công nghệ proteomics không thể khắc phục được các vấn đề trong chuẩn bị mẫu. Các bước như đồng nhất hóa mô, […] - Sự hình thành cầu disulfide trong protein
 Tầm quan trọng của cầu nối Disulfide Ổn định cấu trúc Protein: Cầu nối disulfide giúp ổn định cấu trúc bậc ba và bậc bốn của […]
Tầm quan trọng của cầu nối Disulfide Ổn định cấu trúc Protein: Cầu nối disulfide giúp ổn định cấu trúc bậc ba và bậc bốn của […] - TỔNG QUAN WESTERN BLOTTING
 Western Blotting (WB) là kĩ thuật phân tích protein được sử dụng rộng rãi trong ngành sinh hoá, sinh học phân tử. Là một kĩ thuật […]
Western Blotting (WB) là kĩ thuật phân tích protein được sử dụng rộng rãi trong ngành sinh hoá, sinh học phân tử. Là một kĩ thuật […] - Palmitoyl và khử palmitoyl: Vai trò trong sinh học tế bào và ung thư
 Palmitoyl hóa là quá trình gắn nhóm palmitate vào protein, giúp điều chỉnh vị trí và chức năng của chúng. Quá trình này có thể đảo […]
Palmitoyl hóa là quá trình gắn nhóm palmitate vào protein, giúp điều chỉnh vị trí và chức năng của chúng. Quá trình này có thể đảo […] - Phân tích trình tự kháng thể: Khám phá sự đa dạng và ứng dụng
 Cấu trúc kháng thể Kháng thể, còn được gọi là immunoglobulin, là một cấu trúc hình chữ Y bao gồm bốn chuỗi polypeptide – hai chuỗi […]
Cấu trúc kháng thể Kháng thể, còn được gọi là immunoglobulin, là một cấu trúc hình chữ Y bao gồm bốn chuỗi polypeptide – hai chuỗi […] - Giải trình tự peptide: Công cụ cốt lõi trong nghiên cứu Proteomics
 Giải trình tự peptide là quá trình xác định trình tự các axit amin trong một chuỗi peptide. Đây là kỹ thuật then chốt trong proteomics, […]
Giải trình tự peptide là quá trình xác định trình tự các axit amin trong một chuỗi peptide. Đây là kỹ thuật then chốt trong proteomics, […] - Phân tích axit amin trong dinh dưỡng và công nghiệp thực phẩm
 Axit amin đóng vai trò then chốt trong việc làm sáng tỏ mối quan hệ phức tạp giữa dinh dưỡng và ngành công nghiệp thực phẩm. […]
Axit amin đóng vai trò then chốt trong việc làm sáng tỏ mối quan hệ phức tạp giữa dinh dưỡng và ngành công nghiệp thực phẩm. […] - Phân tích axit amin trong kiểm soát chất lượng protein tái tổ hợp
 Dược phẩm sinh học protein tái tổ hợp Rối loạn chức năng của các protein có trình tự axit amin bất thường hoặc không có […]
Dược phẩm sinh học protein tái tổ hợp Rối loạn chức năng của các protein có trình tự axit amin bất thường hoặc không có […] - TỔNG QUAN VỀ SDS-PAGE
 SDS-PAGE là gì? Điện di trên gel polyacrylamide biến tính với sodium dodecyl sulfate (SDS-PAGE) là một kỹ thuật điện di protein phổ biến trong sinh […]
SDS-PAGE là gì? Điện di trên gel polyacrylamide biến tính với sodium dodecyl sulfate (SDS-PAGE) là một kỹ thuật điện di protein phổ biến trong sinh […] - LIPID HÓA PROTEIN: CƠ CHẾ, PHÁT HIỆN VÀ CÁC BỆNH LÝ LIÊN QUAN
 Protein lipid hóa là gì? Biến đổi sau dịch mã (PTMs) là những thay đổi hóa học xảy ra sau quá trình tổng hợp protein, liên […]
Protein lipid hóa là gì? Biến đổi sau dịch mã (PTMs) là những thay đổi hóa học xảy ra sau quá trình tổng hợp protein, liên […] - Lipid: Nhóm Phân Tử Đa Năng Trong Sinh Học và Công Nghệ
 Lipid là một nhóm lớn các phân tử hữu cơ không phân cực, đặc trưng bởi tính kỵ nước (hydrophobic) và khả năng hòa tan trong […]
Lipid là một nhóm lớn các phân tử hữu cơ không phân cực, đặc trưng bởi tính kỵ nước (hydrophobic) và khả năng hòa tan trong […] - Glycosyl hóa: Biến đổi sau dịch mã và tác động lên cấu trúc chức năng protein
 Con đường đường phân glycosyl hóa (Glycosylation pathway) là một quá trình biến đổi sau dịch mã (PTM) quan trọng, trong đó các gốc glycan được […]
Con đường đường phân glycosyl hóa (Glycosylation pathway) là một quá trình biến đổi sau dịch mã (PTM) quan trọng, trong đó các gốc glycan được […] - Cách mạng hóa Proteomics: Tiến bộ trong công nghệ, tích hợp AI và ứng dụng rộng hơn
 1. Sự phát triển của công nghệ Proteomics 1.1 Proteomics dựa trên khối phổ Proteomics dựa trên khối phổ là một lĩnh vực năng động và then […]
1. Sự phát triển của công nghệ Proteomics 1.1 Proteomics dựa trên khối phổ Proteomics dựa trên khối phổ là một lĩnh vực năng động và then […] - Phản ứng phân hủy Edman
 Phản ứng phân hủy Edman là phương pháp giải trình tự protein được Pehr Edman công bố vào năm 1950. Phương pháp này giúp xác định […]
Phản ứng phân hủy Edman là phương pháp giải trình tự protein được Pehr Edman công bố vào năm 1950. Phương pháp này giúp xác định […] - Applications of Tandem Mass Spectrometry (MS/MS) in Protein Analysis for Biomedical Research
 Applications of Tandem Mass Spectrometry (MS/MS) in Protein Analysis for Biomedical Research 1. Giới thiệu Proteomics – nghiên cứu toàn bộ bộ protein của một hệ […]
Applications of Tandem Mass Spectrometry (MS/MS) in Protein Analysis for Biomedical Research 1. Giới thiệu Proteomics – nghiên cứu toàn bộ bộ protein của một hệ […] - Kỹ thuật định lượng không nhãn
 Kỹ thuật định lượng không nhãn là gì? Ngày nay, các nghiên cứu về proteomics không còn chỉ tập trung vào việc xác định càng nhiều […]
Kỹ thuật định lượng không nhãn là gì? Ngày nay, các nghiên cứu về proteomics không còn chỉ tập trung vào việc xác định càng nhiều […] - Matrix effects and application of matrixeffect factor
 Matrix effects and application of matrix effect factor 1. Thực trạng Hiện nay, LC-MS là một trong những kĩ thuật phân tích tối ưu nhất về […]
Matrix effects and application of matrix effect factor 1. Thực trạng Hiện nay, LC-MS là một trong những kĩ thuật phân tích tối ưu nhất về […]


